A Vernadskian Reconsideration of Galactic Cycles and Evolution


A note to readers: this is an old post on the archive website for Promethean PAC. It was written when we were known as LaRouche PAC, before changing our name to Promethean PAC in April 2024. You can find the latest daily news and updates on www.PrometheanAction.com. Additionally, Promethean PAC has a new website at www.PrometheanPAC.com.
As has been emphasized recently by Lyndon LaRouche and his Executive Intelligence Review magazine and LaRouche Political Action Committee, to understand climate, weather, and the behavior of water on our planet we must start by understanding the role of our galaxy.1
Records of the largest climate variations over the past half billion years correspond to changes in the galactic environment experienced by our Solar System—indicating that the galaxy has the strongest role in determining the climate variations on Earth.2
The implications of this can be looked at in two ways.
On the one side, an adherent to the modern school of scientific reductionism may see this as, perhaps, an interesting phenomenon, but one with no general impact on our understanding of the nature and ordering of causality in the universe.
On the other side, a mind which is not suffering from the debilitating effects of the destruction of science led by David Hilbert and Bertrand Russell3 (mathematical reductionism) will see this as a clue to defining a new understanding of the hierarchical nature of causality in the universe—pursuing the conception of science defined by Nicolas of Cusa (as in his 1440 De Docta Ignorantia) and his follower Johannes Kepler.
Here we will take the opportunity of the publication of the first English translation of Vladimir Vernadsky's 1930 report, “The Study of Life and the New Physics,” to examine another clue, again pointing us towards the need for a higher understanding of our galaxy.4
Studies have shown that there are cycles in the evolutionary development of animal life over the past 540 million years on Earth—cycles which correspond in period and in phase to cyclical aspects of the motion of our Solar System through our galaxy.
This can also be looked at in two ways.
In the modern domination of Russellian reductionism, a “kill mechanism” is sought to explain how different galactic environments can accelerate the extinction rate of species and, thereby, imprint records of these cosmic fluctuations in the evolutionary record.
For an approach freed from the disease of reductionism, we can instead look to the views of Vernadsky, as presented in his 1930 report, “The Study of Life and The New Physics.”
A student of Dmitri Mendeleev, and an avid opponent to the influence of Bertrand Russell on Russian and Soviet science, Vernadsky's hypotheses about life in the cosmos provide an important basis to investigate the relation between the changing expression of life on Earth and the subsuming galactic system.
This provides another avenue for understanding that which subsumes our Solar System, our Earth, and the processes therein.

One of the European Southern Observatory's telescopes in their Very Large Telescope array uses a laser beam to create an artificial star high in the Earth's atmosphere, allowing the astronomers to correct for atmospheric distortion (utilizing adaptive optics) as they study the central regions of our Milky Way Galaxy. The picture was taken in August of 2010 picture by Yuri Beletsky.
Identifying the Important Evidence
Fossil records leave a map of the evolutionary development of complex life on Earth, showing an overall increase in the number of distinct animal species (and more clearly in measures of genera) on the plant over the past 540 million years (as is best recorded in records of ocean life). However, upon this overall increase is imprinted a smaller periodic rise and fall in the number of genera at any given time. Early indication of this go back to the 1980s,5 but more recent analysis (with a more complete fossil record) has solidified the evidence for a cycle in the decline and increase in the number of genera over time.6 Perhaps most interestingly, this cycle corresponds with the period and phase of cyclical aspects of the motion of our Solar System through the Milky Way Galaxy.
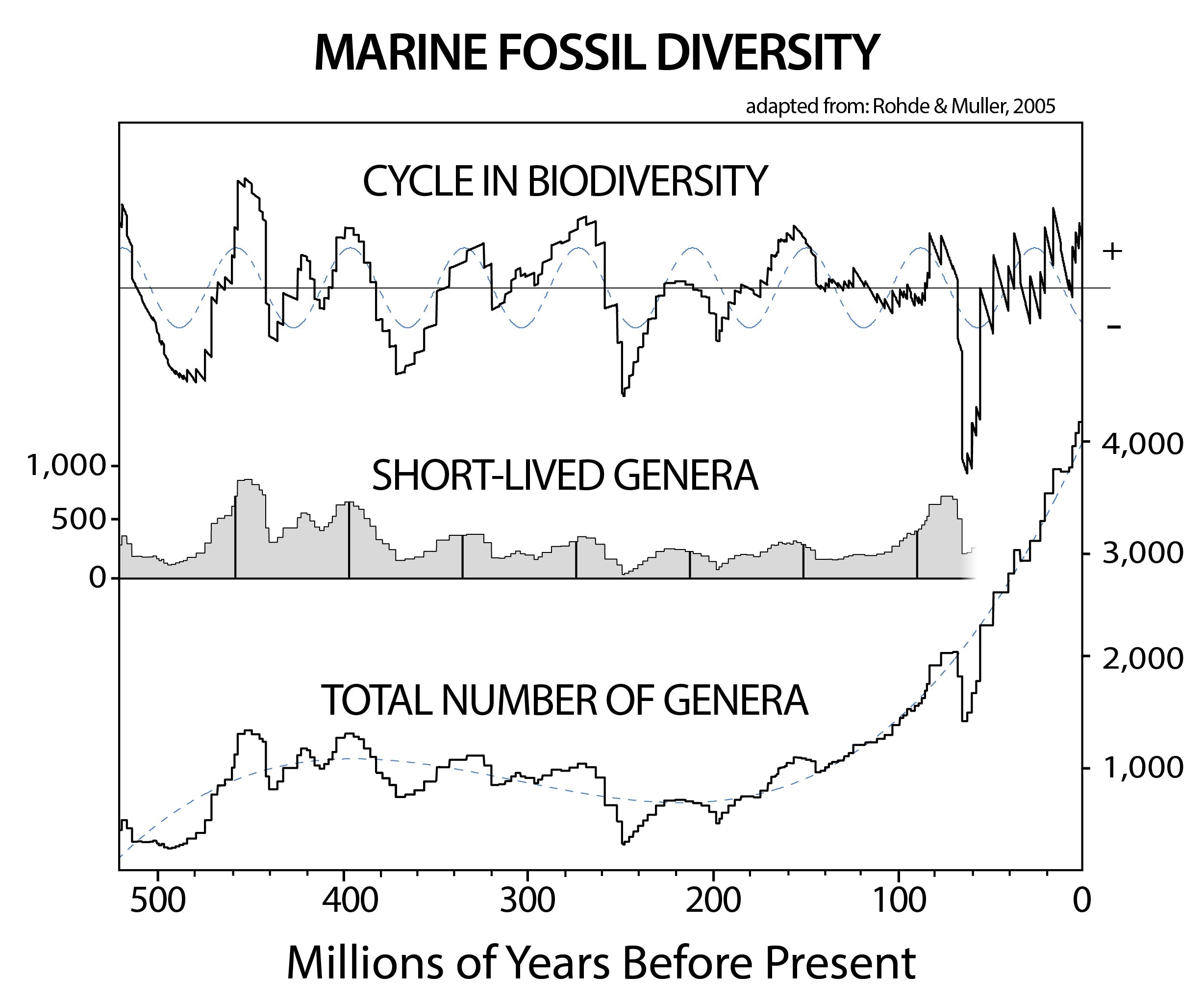
Changes in the total number of fossilized marine genera over the past 540 millions years provides a measure of the changing biodiversity over the Phanerozoic Eon. The total number of distinct genera is shown on the bottom, indicating an overall increase in the biodiversity. When the overall growth trend is removed, the cyclical variation in biodiversity becomes more evident (as shown on the top graph, which includes a 62 million year cycle). If only "short-lived" genera are examined (ones which lasted less than 45 million years) then the 62 million year cycle also presents itself (as shown in the middle graph). The image has been adapted from Rohde and Muller, 2005.
Existing attempts to explain this correlation between galactic activity and evolution of life rely upon a sequence of domino like effects resulting from the introduction of a “kill mechanism.” They look for ways that cosmic processes might kill off large enough numbers of individual animals (either directly, or by creating certain environmental effects which will do so), which in turn could then lead to extinctions of entire species, and, if the killing rate was powerful enough and sustained, then to the extinctions of large numbers of different species, resulting in the extinctions of entire genera, and then families, culminating in a “mass extinction”7
The belief that increased extinction rates, or even mass extinctions can be explained by this type of a bottom-up causality is not a demonstrated generalization based on evidence, but, rather, the product of certain reductionist beliefs and assumptions. In reality the phenomena of mass extinctions are still poorly understood.8 What we know from the fossil record is that there can be relatively rapid (in geological terms) transitions where many species, genera, and families disappear from the records and are replaced by new forms (though these more dramatic (and rapid) shifts exist within the context of an already ongoing slower turnover rate). How and why this occurred the way it did is still not well understood.

Fossilized remains of an extinct species of sea stars (Dipsacaster africanus) from around 130 million years ago. The fossils were discovered in Taba, Morocco. Credit: Didier Descouens
So, rather than assuming we must accept a reductionist framework, here we will take a different approach.
Perhaps most important for this shift in approach is to recognize that it isn't simply extinctions which define these cycles, but extinctions and originations (the generation of new species, genera, and families).
As stated in a 2013 paper on the subject, the evidence for a cycle in the process of the evolutionary development of life on Earth “results from the coherent interaction of both extinction and origination fluctuations, producing a stronger signal than either would or could alone”.9
So we must also ask why there exist periodic phases characterized by the origination of new genera.
Put simply, we're looking for more than a kill mechanism, we're examining, on the one side, the anti-entropic development of life on Earth, and, on the other, the relation of our Solar System to our galactic system—and we're asking why cycles in both processes correlate so well. The work of Vernadsky provides a new basis to investigate this relation, in these top-down terms.
Vernadsky's “The Study of Life and the New Physics”
We don't know what life is.
Vernadsky's work provides an important distinction between the study of living processes and life per se. We can study living processes as effects of life, as particular expressions of life, without assuming that these specific expressions, alone, define life per se. This important distinction provides the needed framework to properly pursue the properties and characteristics of life, per se—investigating that which underlies certain particular expressions and manifestations.
Vernadsky took up exactly this approach in his 1930 report, “The Study of Life and the New Physics.” Examining the identifiable properties of living processes—as they can be studied in the context of their existence in the biogeochemical medium of the Earth's biosphere—he separated the properties into two lists: first, those properties which are associated with the planetary (biogeochemical) medium within which living processes are manifested on Earth; second, those properties displayed by living processes which can not be attributed to the characteristics and properties of this planetary context, and, thus, might express something more universal about life, per se.10
Vernadsky immediately follows this second list with a conclusion which will be upsetting to today's reductionists: “This list is not complete, but it indicates, with evidence, that life manifests itself in the Cosmos in other forms than those which biology normally displays.”
Since living processes are not merely a phenomenon of geochemistry11—but are an expression of a principle of life, per se, manifested in the context of a geochemical medium—we should be willing to seek out in the cosmos other expressions of these non-planetary properties of life.
Vernadsky then dedicates the entire latter half of his report to the two non-planetary properties of life which he thinks could be the most fruitful in investigating how “life manifests itself in the Cosmos in other forms than those which biology normally displays.”
Here I will dwell upon two phenomena which will allow for the clarification of the important role which the investigation of life plays in the scientific picture of the Universe, created by the new physics, notably upon the dissymmetry of the space of living organisms and on biological time. In the first case, this is a matter of new properties (a particular state of physical space), observed in living organisms, and in the second, new properties of physical time.12
Of his 18 section report, Vernadsky focuses most of the latter half to the first of these two, “the dissymmetry of the space of living organisms” (sections 11 to 16), followed by one section on biological time (section 17).
Vernadsky's work—both distinguishing a principle of life, per se, from the particular expressions of living processes we're familiar with on Earth, and positing the need to investigate other potential expressions of this principle in the cosmos—provides a critical, non-reductionist basis for investigating the correlation of cycles of extinction and origination in the fossil record with the cycles of our Solar System's motion through our galaxy—that is, to investigate the potential relationship between the process of the anti-entropic development of living processes on Earth and the processes of the cosmic system of our galaxy.
As we will see, Vernadsky's conception of dissymmetrical states of space will be key.
Cosmic Dissymmetry
In a different address (delivered one year later), Vernadsky made some rather interesting remarks regarding galactic systems specifically. Citing early studies examining the distribution of “spiral nebulae” (as spiral galaxies used to be called), Vernadsky hypothesized their orientations could be an expression of a “dissymmetrical” characteristic of the cosmos.
The spiral form of nebulae and of some stellar agglomerations indicates the probable presence of analogous dissymmetrical phenomena in the Cosmos. If the right spirals predominate in effect, clearly, among the spiral nebulae, as numerous photographs attest, or in certain parts of the universe right spiral nebulae are concentrated and in others left spiral nebulae, the existence of dissymmetric spaces in the Cosmos would become more than probable. This dissymmetry would seem to be analogous to that which we observe in the space penetrated by life, that is to say, that it possesses enantiomorphic vectors and both of the vectors—left and right—could exist there at the same time, but not in equal number; the right-handed vectors most often predominate there.13
While recent studies indicate Vernadsky may have been onto something interesting regarding the large-scale distribution of galaxies,14 here we're interested in the potential dissymmetrical characteristics of a single galaxy—our own.
For a single spiral galaxy to express and inherent dissymmetry—i.e. to have an inherent handedness – there has to be a physical distinction between the top and bottom (north and south),15 a distinction expressing the global characteristics of the galactic system as a whole.
Most importantly, if we are working from Vernadsky's conception of potential cosmic expressions of a quality of dissymmetrical space which we see expressed in living organisms, then perhaps the top-bottom (north-south) distinction which defines the dissymmetry of a spiral galaxy should be expressed in the response of living processes most strongly. That is, it would make sense that the most important evidence for defining an inherently dissymmetrical space of a galaxy would be the reaction of living processes to the influence of that dissymmetrical space.
Holding that thought, let's return to what we know about the relationship of our Solar System to the galaxy.
As we orbit around the center of our galaxy, the Solar System also passes above and below the galactic plane, in a bobbing-type motion. Based on current measurements and analysis, the cycles of this up and down motion are roughly 30 (26-37) million years from mid-plane, through a peak, back to mid-plane, or 30 million years from one peak to the opposite peak, or 60 million years from one peak, through the opposite, and back to the same side.
Most researchers think that the conditions on either side of the galactic plane (north or south) should be generally similar, and, therefore, any imprint of this changing galactic environment recorded in the Earth's history should express a 30 million year periodicity.
In fact this is true for at least one abiotic process, the climate, where a 30 million year cycle has been found.16
However, records of the evolutionary development of life on Earth display a ~62 million year fluctuation.17 As mentioned above, this biodiversity cycle is strongest when not only examining extinctions, but extinctions together with originations (the appearance of new genera), forcing the investigation beyond the reductionist search for a kill mechanism.
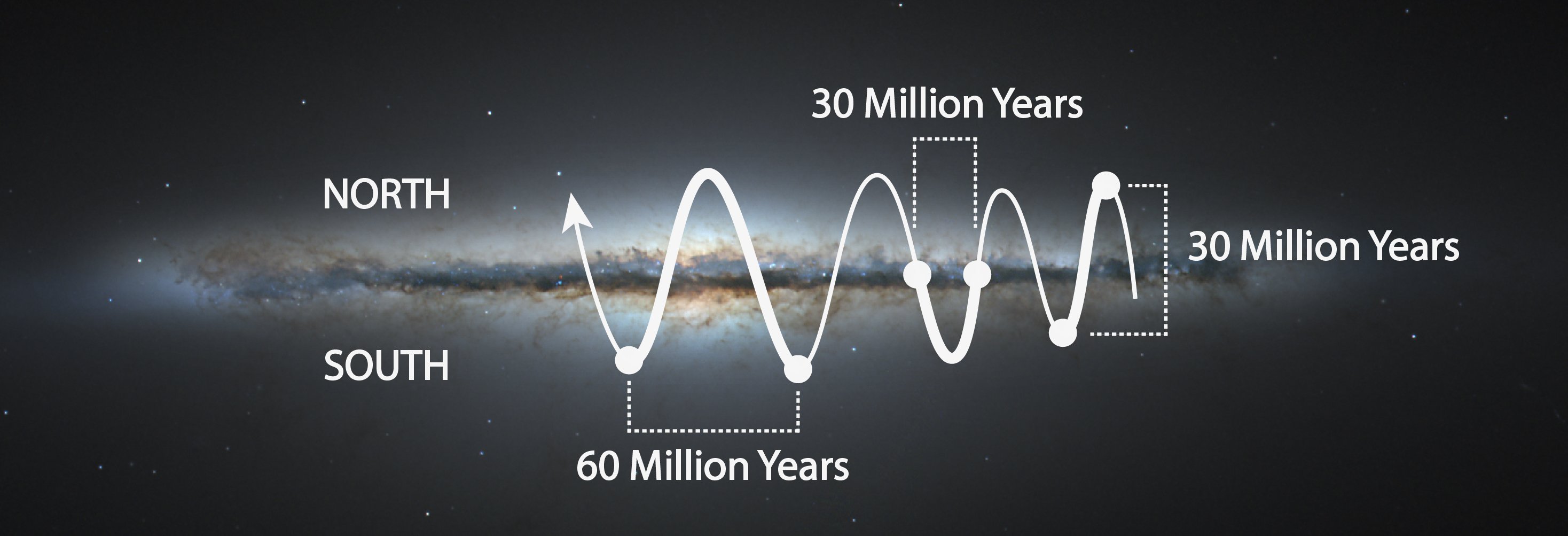
An illustration of the motion of our Solar System above and below the galactic plane (original image of NC galaxy 5866 from NASA, ESA, and The Hubble Heritage Team (STScI/AURA)).
Thus, the evidence for a relationship between processes of our galactic system and the evolutionary development of life on Earth is not simply associated with being either above or below the galactic plane, but with the characteristics of one side vs the other. From within the reductionist camp this is taken as evidence to doubt the existence of a connection between this galactic process and the evolution of living processes on Earth (despite the clear correlation), because they have no reason to hypothesize a distinction between the north and south sides.18 But when viewed from the conceptions of Vernadsky, the distinction which serves as their basis for doubt becomes our point of interest.
A physical distinction between one side of the galaxy from the other is required for our Vernadskian hypothesis of a dissymmetrical characteristic governing the physical space of the galactic system – providing the critical evidence needed to define a distinct, intrinsic handedness of the system (irrespective of one's vantage point).
The evolutionary cycle being 60 million years, rather than 30 (and matching the proper phase), provides the needed evidence for a distinction, indicating the potential for an inherent difference in the north vs south sides of our galaxy, and, thereby, its inherent dissymmetry. It is most appropriate that fluctuations in the history of the evolutionary development of living processes on Earth are what provides the critical evidence for defining an intrinsic dissymmetry of our galactic system—indicating galactic manifestation of dissymmetrical space, to which living processes on Earth are responsive.19
Spacetime of Anti-Entropy
In the terminology and framework pursued by Vernadsky, this could be an expression of a dissymmetrical spacetime characteristics of our galactic system.20
This is not the first indication that the study of galactic systems could require a new conception of a self-bounded spacetime intrinsic to that galactic system.21 However, Vernadsky's direction of work indicates that we should open our minds to the qualities of the spacetime characteristics of living processes (rather than simply abiotic physics), if we are to truly attempt to understand the cosmos as containing a principle of life, per se, and galactic systems therein.
With this evidence for a relation between the evolutionary development of life on Earth and the processes of our galactic system, we see the option to invert the investigation—to examine the characteristics expressed by evolution as informing us about the nature of our galactic system as a whole.
As Vernadsky correctly identified in his 1926 address on evolution,22 there is an intrinsic direction in the evolutionary development of life on Earth—the increasing energy flux density of the biosphere system—which Vernadsky called his “second biogeochemical principle.”
This biogeochemical principle which I will call the second biogeochemical principle can be formulated thus: The evolution of species, leading to the creation of new, stable, living forms, must move in the direction of an increasing of the biogenic migration of atoms in the biosphere…
[This second biogeochemical principle] indicates, in my opinion, with an infallible logic, the existence of a determined direction, in the sense of how the processes of evolution must necessarily take place… All theories of evolution must take into consideration the existence of this determined direction of the process of evolution, which, with the subsequent developments in science, will be able to be numerically evaluated. It seems impossible to me, for several reasons, to speak of evolutionary theories without taking into account the fundamental question of the existence of a determined direction, invariable in the processes of evolution, in the course of all the geological epochs. Taken together, the annals of paleontology do not show the character of a chaotic upheaval, sometimes in one direction, sometimes in another, but of phenomena, for which the development is carried out in a determined manner, always in the same direction, in that of the increasing of consciousness, of thought, and of the creation of forms augmenting the action of life on the ambient environment.23
Since Vernadsky's time, we've accumulated a much larger and more detailed map of the evolutionary development of life. While the new evidence strongly conforms to Vernadsky's second biogeochemical principle,24 we are still far from understanding the principle which has composed that map.
In pursuit of this, we've been pointed to the processes of our own galactic system—as the macroevolutionary pulsations associated with the anti-entropic development of living processes on Earth beat in harmony with our Solar System's experience of the dissymmetrical characteristics of our galaxy.
Rather than simply an Earth-based phenomenon, the development of life on Earth could be an expression of an anti-entropic character of our galaxy, returning us to the opening challenge: understanding the causal role of our galactic system in the hierarchical ordering of the Universe.